|
|
|
L’anatomie et les fonctions des cellules
Introduction (I) Pour bien comprendre ce qu'est la vie et toute sa complexité, nous devons nous pencher sur son entité la plus simple, la cellule à vrai noyau, dite eucaryote, pour tenter de comprendre comment elle fonctionne et de quelle manière elle réagit face à son environnement. Se greffe sur cette faculté, l'interaction de l'organisme tout entier avec la biosphère, un sujet qui concerne plus l'écologie et dans une certaine mesure la thermodynamique, autant de thèmes sur lesquels nous nous attarderons dans d'autres dossiers. Nous allons aborder la question du fonctionnement des cellules, principalement eucaryotes (à noyau et organites), à travers l'étude de l'ADN, la structure cellulaire et de ses facultés de reproduction, avant d'aborder la question des parasites et clôturer cet article sur quelques questions en suspens[1]. Le corps humain : un organisme autonome Selon une étude publiée en 2016 par Ron Senders de l'Institut des Sciences Weizmann d'Israël et ses collègues, le corps humain (un homme de 20-30 ans, pesant 70 kg pour 1.70 m) est constitué d'environ 30 trillions soit 30000 milliards de cellules (ce qui représente ~1028 atomes) qu'on peut regrouper en quelque 200 types de cellules spécialisées allant de la taille l'oeuf à celle du neurone ou du globule rouge qui formeront la vingtaine de tissus différents que comprend le corps humain. Le cerveau d'un adulte contient environ 86 milliards de neurones (et autant de cellules non neuronales) et 100 milliards de cellules forment l'épiderme. Toutes ces cellules sont organisées dans un but commun : maintenir l'organisme en bon état. Comme une machine, nos tissus et nos organes s'usent et nos fonctions métaboliques peuvent se dérégler. Mais contrairement à une machine, du fait que nos organes sont vivants, des mécanismes nous maintiennent en permanence dans le meilleur état possible, mieux encore que la machine la mieux entretenue. Pour assurer cette fonction vitale, 300 millions de cellules meurent toutes les minutes et 300 millions sont renouvellées ou créées chaque jour, du plus profond de nos entrailles à la surface de notre épiderme. Comme tous les organismes vivants, le corps humain est non seulement autonome et ne compte que sur sa propre énergie pour assurer ses fonctions mais c'est un organisme à la fois stable et instable, ce qui est une condition étrange et a priori paradoxale mais primordiale pour garantir son évolution. En effet, tout organisme eucaryote doit être stable du point de vue biologique, c'est-à-dire qu'il doit conserver ses caractéristiques au fil des générations. Mais d'un autre côté, sur le plan thermodynamique, un être vivant doit être instable afin d'être capable de réagir et de s'adapter à son environnement. La plupart du temps il s'agit de fonctions réflexes héritées d'un lointain passé où nous n'étions même pas encore humains et aujourd'hui profondément enfouies dans l'ADN de chaque cellule. Si l'homme était dans un état thermodynamique stable, en d'autres termes s'il prenait à notre corps l'envie de ne plus s'adapter, de ne plus réagir aux solicitations nerveuses, aux substances chimiques, de ne plus évacuer les déchets, de ne plus réguler la respiration et la circulation sanguine en fonction de l'activité cellulaire, bref de ne plus rien faire, ce serait l'anarchie en quelque minutes : suffocation, douleurs cardiaques, nausées, paralysie, bref la mort de l'organisme.
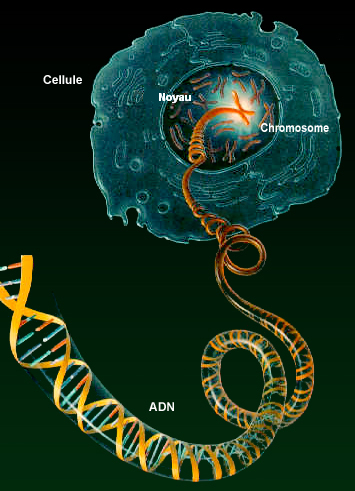
Ainsi, contrairement à une machine qui doit fonctionner de la manière la plus stable possible, le corps humain doit être thermodynamiquement instable pour assurer ses fonctions. La machine à toutes les chances de s'user et de tomber en panne, tandis que le corps humain bénéficie d'une assurance-vie assortie d'un dépannage discret à perpétuité, ou du moins valable environ un siècle. Cette faculté d'adaptation extraordinaire qui nous suit durant la vie nous donne déjà une idée du degré de complexité des organismes évolués mais malheureusement personne ne comprend très bien comment tous ces éléments interagissent et évoluent. Déjà au stade du foetus, c'est la grande inconnue. L'embryon se métamorphose mais personne ne sait vraiment par quelle magie ! On peut donc supposer qu'au centre de chaque cellule préside un "centre de contrôle relais" qui assure le transit des informations et veille localement à la bonne exécution des tâches. Il semble respecter un plan de construction ou de maintenance général mais que personne n'a encore trouvé. Mais peut-être ne cherchons-nous pas au bon endroit ou l'arbre cache-t-il peut-être la forêt... Il s'agit toutefois d'une vision mécaniste de notre métabolisme qui reste très naïve. Ainsi que nous l'avons expliqué à propos de la chimie prébiotique, lors des balbutiements des premières formes de vie dans les marais chauds de Darwin, de toute évidence elles ne disposaient pas encore de "centre de contrôle relais" ni même de centre de contrôle du tout et pourtant ces créatures ont survécu et même évolué ! Mais le développement le plus abouti fut l'émergence des cellules eucaryotes disposant d'un véritable noyau. On y reviendra. L'ADN contenu dans le noyau cellulaire, car c'est bien de lui qu'il s'agit, est une structure apparue très tard dans l'histoire de la vie, il y a environ 1.2 milliard d'années, puis seulement sont apparus les vertébrés (500 millions d'années) ensuite les mammifères (200 millions d'années) pour aboutir à l'homme vieux d'à peine 7 millions d'années en tenant compte de notre ancêtre Toumaï. Qu'a donc l'ADN de si particulier pour avoir été intégré dans tous les organismes vivants et même les virus ? Tentons de répondre à cette question et à quelques autres. Le code génétique On savait depuis la fin du XIXe siècle que certains gènes contribuent au développement embryonnaire des végétaux et des animaux. On avait même observé que leur mutation entrainent parfois des chimères. C'est en 1902 que le médecin britannique Archibald Edward Garrod établit un lien entre une enzyme et un gène. Mais il faudra attendre 1958 pour que Jérôme Lejeune décrive la trisomie 21, une anomalie génétique qui frappe le chromosome 21 et à l'origine d'une déficience intellectuelle d'origine génétique. Ensuite, après que Watson et Crick aient réussi à décrire la structure de l'ADN en 1953, un grand pas fut franchi en 1963-64 lorsque Nirenberg, Khorana et Ochoa sont parvenus à déchiffrer le code génétique, c'est-à-dire la correspondance entre l'ADN composé des nucléotides ATCG et l'expression des gènes codant les polypeptides constitués de 20 acides aminés différents. A partir de ce moment là, les généticiens disposaient des bases de la biologie moléculaire et comprenaient enfin le génome et comment son contenu était traduit en protéines. On reviendra sur ces notions. Au sein d'une même espèce, les individus présentent à la fois des caractères communs et des caractères individuels. Ce paradoxe apparent s'explique par les propriétés du code génétique, l'ADN. En effet, l'ADN ou acide désoxyribonucléique est regroupé en gènes porteurs de l'information héréditaire; c'est le génome. Mémoire génétique de l'organisme, il est présent dans le noyau de chacune des cellules de notre corps, des neurones du cerveau à l'épiderme, en passant par les gamètes. A ne pas confondre avec d'autres structures tels que les poils ou les ongles qui sont constitués de protéines (kératine), des substances sur lesquelles nous reviendrons.
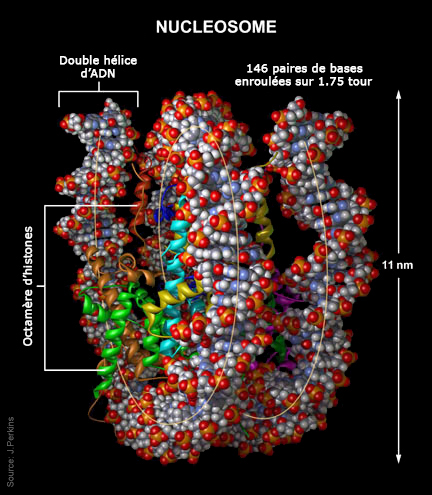
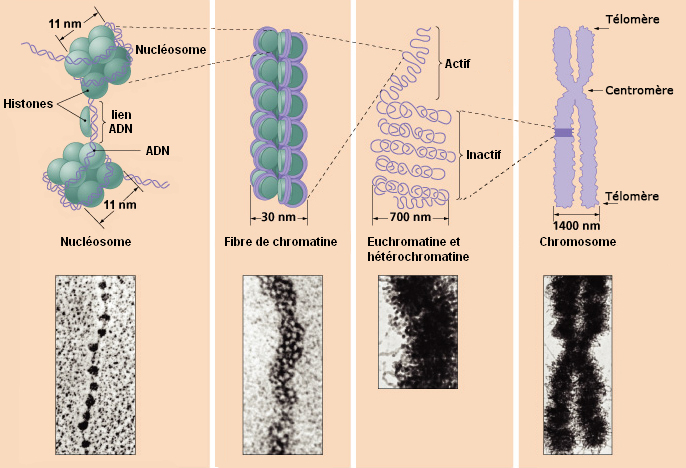
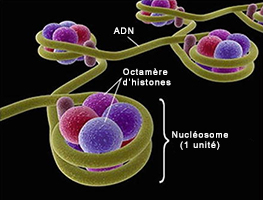
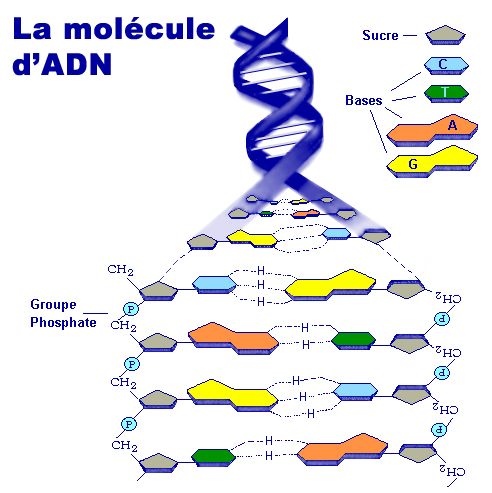
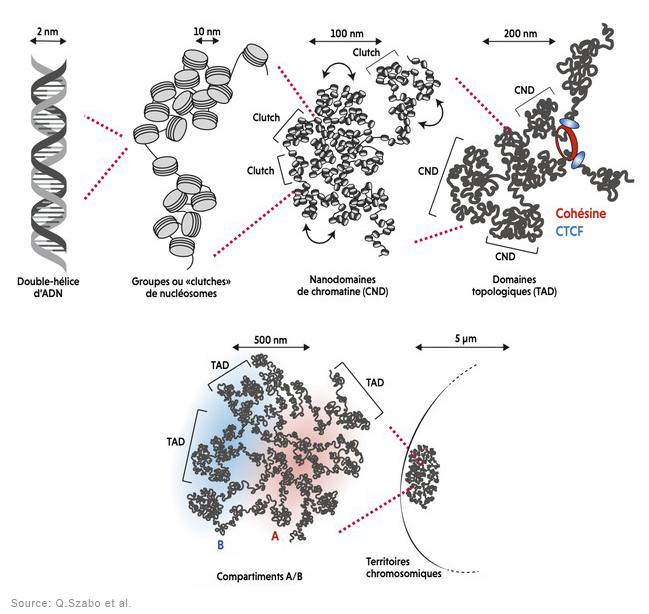
La molécule d'ADN étant tellement grosse tant en masse qu'en volume (voir plus bas) que la nature l'a compactée au maximum et dotée de capacités étonnantes afin que la machinerie cellulaire puisse la manipuler plus facilement, notamment pour la synthèse des protéines et la reproduction sur lesquelles nous reviendrons. A l'échelle macroscopique, la molécule d'ADN enfouie au coeur des chromosomes présente une succession de pliages et de compactions d'une rare complexité qui, ajoutée à sa faculté de mémorisation, rend la molécule d'ADN unique en son genre. La molécule d'ADN qui mesure quelques centaines de nanomètres de longueur a dut subir un processus de compaction d'un facteur 7 au niveau nuléosomique (le plus élémentaire), puis un enroulement sous forme de fibres de chromatines d'un facteur 6 afin de tenir dans le volume minuscule du chromosome ! Voyons ce mécanisme en détail et profitons-en pour définir ses principaux composants. Comme l'explique le schéma ci-dessus, l'ADN présente une organisation tridimensionnelle très élaborée subdivisée en plusieurs niveaux ou échelons. Au niveau fondamental se trouve le nucléosome, dont la structure cristallographique fut mise en évidence en 1997 (cf. T.J. Richmond et al.). Chaque nucléosome est organisé dans des unités répétitives de 11 nm comprenant 8 molécules d'histones (des protéines basiques riches en lysine et arginine) appelées un octamère d'histones autour duquel s'enroule l'ADN sur un tour trois-quart sur une longueur de 146 paires de bases ou nucléotides. Ces nucléosomes sont reliés entre eux par une molécule d'ADN de 2.5 nm de diamètre appelée le lien internucléosomique. A lire : How DNA Is Packed In The Cell, Brian D. Strahl/UNC, 2014
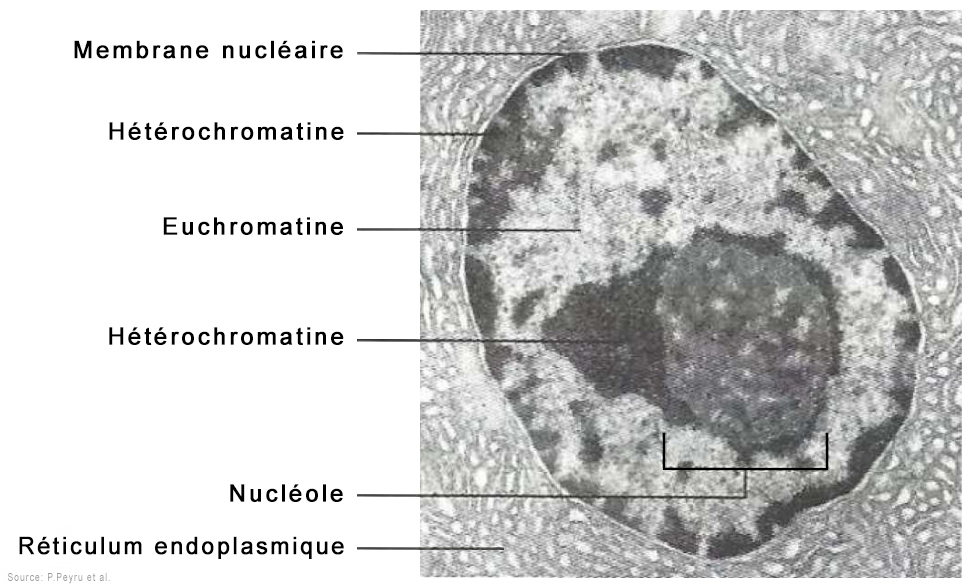
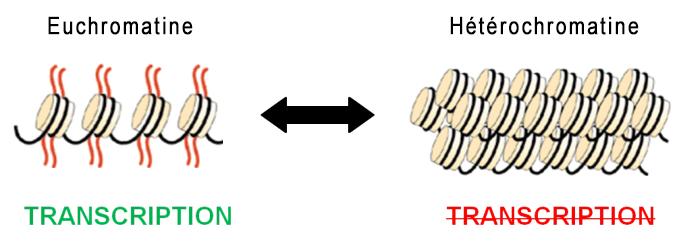
Ces chapelets de nucléosomes sont ensuite rassemblés dans une fibre de chromatine d'environ 30 nm de diamètre et excessivement longue comprenant l'ADN, l'ARN et les histones. Ces chromatines existent sous deux états, l'une active appelée l'euchromatine, l'autre inactive appelée l'hétérochromatine. Cette différence est essentielle. L'euchromatine ou plus précisément les histones de l'euchromatine correspondent à une chromatine décompactée et déspiralisée correspondant aux zones actives des gènes. Elles sont accessibles aux ARN polymérases réalisant la transcription. L'euchromatine active est principalement localisée à l'intérieur du nucléoplasme et représente environ 10% de l'ADN, 80% du restant étant sous une forme inactive hybride entre l'euchromatine et l'hétérochromatine. Le second état de la chromatine est représenté par les histones de l'hétérochromatine. Elles correspondent à une chromatine condensée et donc inactive où généralement les gènes ne sont pas exprimés. Elles ne sont pas accessibles aux ARN polymérases (la transcription est inactive) mais localement des régions sont transcrites. L'hétérochromatine est généralement localisée en périphérie du noyau et du nucléole. Ce type de chromatine représente 10% de l'ADN et se trouve généralement près des centromères et des télomères des chromosomes.
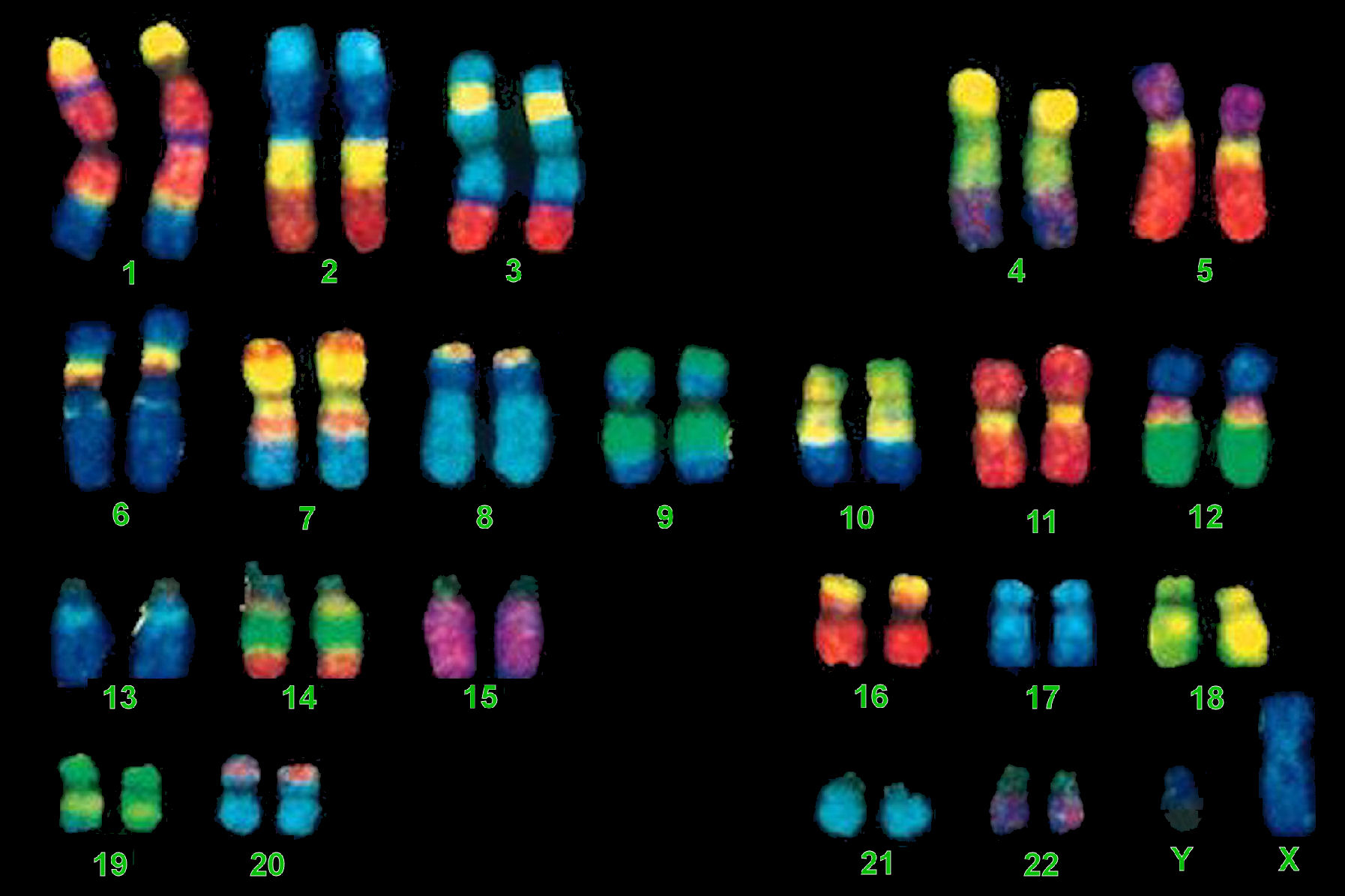
Ces fibres de chromatines mesurent environ 700 nm de diamètre. Etant donné qu'elles sont très longues, elles sont enroulées sur elles-mêmes puis hautement compactées dans les chromosomes dont la largeur est d'environ 1700 nm ou 0.0017 mm soit 100 fois plus fin que l'épaisseur d'un cheveux (~0.1 mm ou 100 microns). Vu de l'extérieur, chaque chromosome dispose d'un région plus ou moins médiane pincée appelée centromère qui intervient lors de la division cellulaire. Aux extrémités du chromosome se situent les télomères qui déterminent la fin des séquences génétiques et évitent aux chromosomes de fusionner entre eux. Récemment, on a découvert que ces télomères contenaient de l'ARN qui peut jouer un rôle dans le vieillissement cellulaire puisque les télomères raccourcissent à chaque division cellulaire. Le génome humain comprend 46 chromosomes répartis en 23 paires : 22 paires de chromosomes homologues ou autosomes plus 1 paire de chromosomes dits sexuels ou gonosomes (hétérochromosomes), XX chez la femme et XY chez l'homme[2]. On reviendra sur le chromosome Y dans l'article consacré à l'avenir de l'Homme. Les 22 paires de chromosomes autosomes sont numérotés de 1 à 22 et classés en 7 groupes de A à G en fonction de leur taille et de la position du centromère. Cet ensemble de chromosomes forme le caryotype (l'arrangement du génome) et varie d'une espèce à l'autre. A
voir : Les acides nucléiques,
R.Querant
Notons que la taille du génome, c'est-à-dire le nombre de paires de chromosomes (la valeur C en génétique) n'est pas représentative de la complexité de l'organisme et varie d'une espèce à l'autre. A ce jeu, l'être humain n'est pas au sommet de cette échelle. Prenons quelques exemples. La fourmi mâle australienne Myrmecia pilosula possède 1 chromosome, le champignon Penicillum de 4 chromosomes, la mouche drosophile de 8 chromosomes, le maïs de 20 chromosomes, la pomme de 34 chromosomes, le chat de 38 chromosomes, la souris de 40 chromosomes, le grand dauphin de 44 chromosomes, le cheval de 64 chromosomes, le chien de 78 chromosomes, le poisson rouge de 100 chromosomes et le papillon Azuré de l'Atlas de ~440 chromosomes. Le record est détenu par la fougère Ophioglosse réticulée qui possède 1440 chromosomes. La molécule d'ADN
A l'échelle moléculaire, le génome humain, l'ADN, comprend quelque 3.055 milliards de paires de bases nucléiques ou nucléotides et 19969 gènes distribués dans 23 paires de chromosomes. Ces quelque 3 milliards de paires de bases représentent une macromolécule de plus de 12 milliards d'atomes ! Par comparaison, la mouche dispose de 117 millions de paires de bases, le maïs de 2 milliards de paires de bases, l'axolotl de 28.4 milliards de paires de bases et la salamandre de 34 milliards de paires de bases. Soulignons qu'il s'agit bien de "paires de bases" (pb) car les bases nucléiques de l'ADN sont appariées par deux à leur bases complémentaires le long de deux brins complémentaires d'ADN, c'est la fameuse double hélice inversée (composées de deux séries antiparallèles de nucléotides en vis-à-vis). On dit que l'ADN est bicaténaire ou double brin. En revanche, pour l'ARN on ne parle pas de paires car il n'y a qu'un seul brin (une seule hélice ou brin torsadé); l'ARN est monocaténaire ou simple brin. L'ARN humain comprend 81 millions de bases (contre ~30000 bases pour le SARS-CoV-2), y compris des séquences non-codantes (introns) mais qui sont supprimées lors de la copie en ADNc. Comme illustré ci-dessous, chaque maillon de cette macromolécule d'ADN contient quatre bases azotées ou nucléotides : l'adénine (A), la thymine (T), la guanine (G), la cytosine (C) complété par un sucre simple (S) et un acide phosphorique (P), le phosphodiester qui joue le rôle d'armature de la double hélice. Les bases azotées s'apparient toujours sous la forme C-G et A-T reliées par 2 ou 3 liaisons hydrogène selon la molécule. La répétition de ces 4 lettres A, T, G et C dans un ordre précis forme le matériel génétique unique de chaque individu. Rappelons que la modification (suppression ou substitution) d'un seul atome ou d'une base de cette chaîne nucléique (mais plus généralement la modification de plusieurs bases) peut avoir un impact sur le corps humain, en particulier sur la santé. Ainsi, comme le variant d'un virus peut augmenter sa transmissibilité et son pouvoir infectieux, un variant de gène peut déclencher des maladies infectieuses, parfois héréditaires. Dans le cadre du projet 1000 Genomes Project (2008-2015), des chercheurs ont découvert 2 millions de variants de gènes dont seulement 622 sont connus sur le plan médical. A
voir : Extraction
de l'ADN de la salive A expérimenter : Extraction de l'ADN d'une banane
La double hélice de l'ADN peut prendre 3 configurations ou formes selon la séquence et la concentration ionique du milieu : - Forme A ou ADN-a : hélice à droite, les paires de bases sont inclinées de 19° par rapport au plan perpendiculaire à l'axe de l'hélice, le grand sillon est bien visible tandis que le petit sillon est écrasé et indiscernable; - Forme B ou ADN-b : hélice à droite, les paires de bases sont perpendiculaires au plan de l'axe de l'hélice, le grand et le petit sillons sont bien visibles (comme ci-dessous à droite). C'est la forme la plus courante; - Forme C ou ADN-z : hélice à gauche, les paires de bases sont perpendiculaires au plan de l'axe de l'hélice, les deux sillons sont équivalents. Elle est assez rare. Dix paires de nucléotides (exactement 10.5 pb) ou paires de bases forment une spire ou un pas hélical de 3.6 nm (3.6 millionièmes de millimètre) de longueur. L'extension complète des 3 milliards de paires de bases de l'ADN humain forme un fil d'environ 1.27 m de longueur et de 2 nm de diamètre qui tient à l'aise dans un centimètre cube au fond d’une éprouvette ! Pour l'anecdote et bien comprendre combien cette machinerie génétique est longue, si nous attachions l'ensemble des brins d'ADN de toutes nos cellules soit au moins 1 trillion de paires de bases (voire 37.2 fois plus selon les estimations), on pourrait faire au moins 600 fois l'aller-retour Terre-Soleil ! Si vous tapiez les lettres du génome humain à raison de 60 mots par minute pendant 8 heures par jour, il vous faudrait entre... 50 et 1860 ans pour le transcrire. Enfin, si les 3 milliards de paires de bases étaient superposées en laissant l'épaisseur d'une feuille de papier (0.1 mm) entre chacune, la pile atteindrait 787 fois la hauteur de l'Empire Empire State Building (381 m) soit 300 km d'altitude ! A consulter : Base de données du génome, EMBL-EBI GeneCards, Compendium du génome humain A lire : 23andMe décode votre ADN (sur le blog, 2007)
La longueur de l'ADN ne reflète pas le degré d'évolution. Plus que la dimension, c'est le contenu, la fonction des gènes, qui est essentielle. La longueur de l’ADN des oiseaux atteint 30 cm et celle de la grenouille peut atteindre 2.40 m ! Chez l'être humain, si l'information du génome était transcrite sur papier, cela représenterait une encyclopédie de 500 volumes de 800 pages chacun mais dont plus de 97% du contenu soit 485 volumes ne codent aucune protéine et n'ont donc apparemment plus d'utilité car les gènes ne sont plus exprimés. Mais nous verrons que cela ne veut pas dire qu'ils soient morts ou ne jouent aucun rôle comme les généticiens l'ont tout d'abord cru. On y reviendra à propos des gènes de rebut. Séquençage du génome et cartographie 3D des chromosomes Le génome humain fut séquencé en grande partie dans le cadre de l'initiative HUGO (Human Genome Organisation) entre 1990 et 2003 et les quelque 8% manquants furent séquencés par les membres du consortium Telomere to Telometre (T2T) entre 2013 et 2022. La version intégrale est nommé T2T-CHM13v2.0 (cf. S.Nurk et al., 2021; Nature, 2021; S.Nurk et al., 2022). On reviendra sur le séquençage du génome à propos du génie génétique. Grâce aux progrès réalisés en biologie moléculaire comme la méthode Hi-C (ou HiC-seq) de séquençage à haut débit combinés aux progrès en informatique, les chercheurs ont pu cartographier les chromosomes en 3D, dont les différentes échelles d'organisation de la chromatine qu'ils contiennent. Ils ont découvert que la chromatine s'organise sous forme d'une succession de boucles et domaines compartimentés. Les chercheurs ont ensuite analysé la fréquence des contacts entre différentes domaines génomiques (cf. J.Dekker et al., 2009) pour tenter de découvrir des motifs inhabituels et ont étudié la conformation spatiale de la chromatine et des domaines topologiques associés ou TAD (cf. B.Ren et al., 2012; S. Razin et al., 2016; F.Bantignies et al., 2019), conduisant à de multiples découvertes.
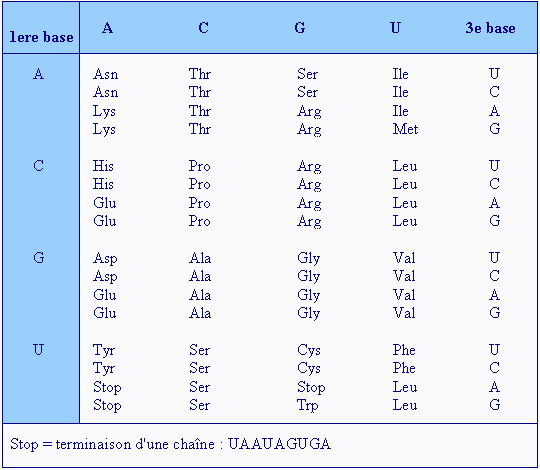
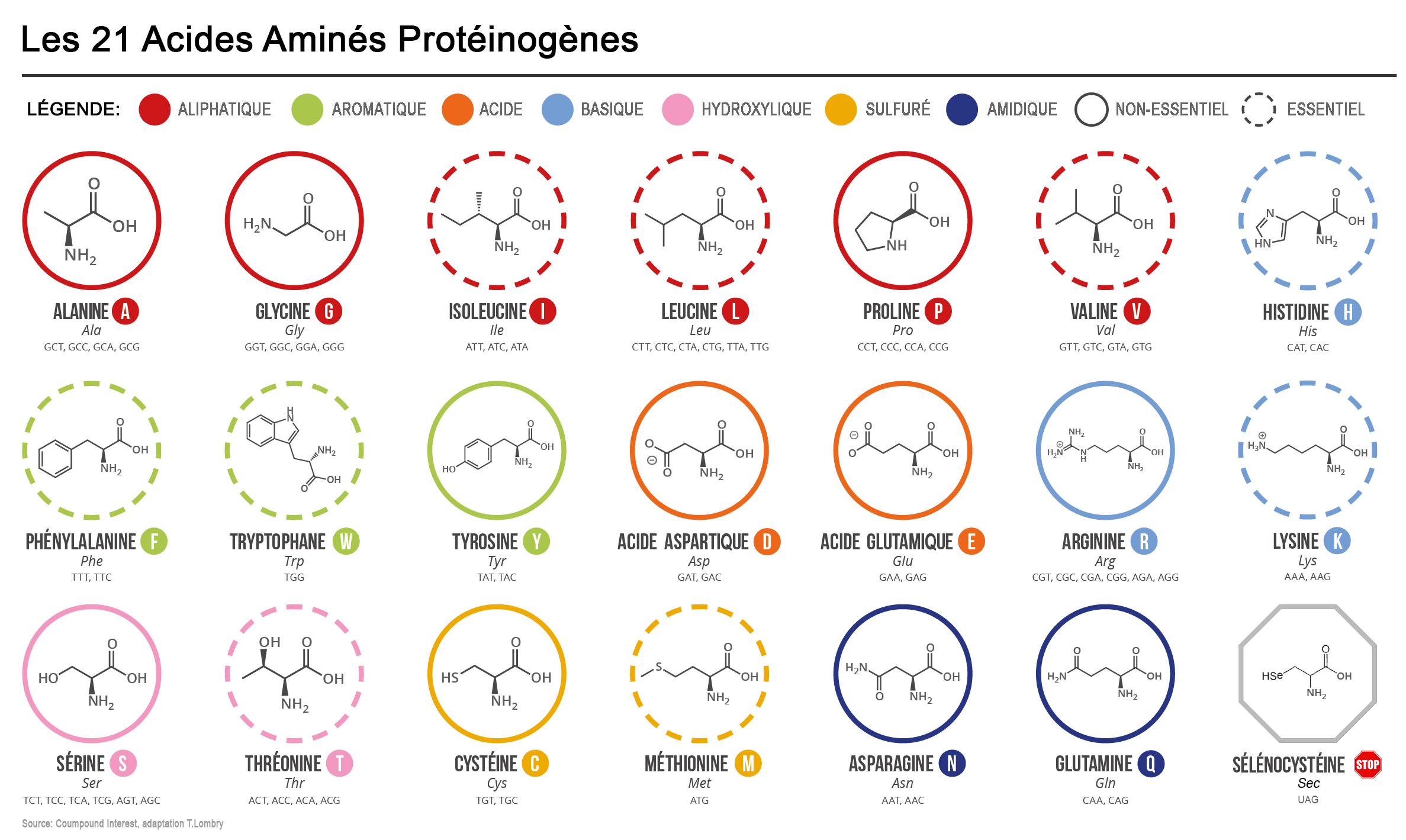
Qu'est-ce qu'un TAD ? Un TAD (l'acronyme de Topological Associated Domain) est un domaine génomique, une région compartimentée de la chromatine caractérisée par des interactions internes. Le génome est ainsi divisé en une série de TAD. Les TAD se rassemblent préférentiellement avec d'autres TAD de même type pour former des architectures tridimensionnelles ordonnées appelées territoires chromosomiques. Plus intéressant, les TAD sont également des régions de réplication de l'ADN et de régulation de l'expression des gènes (cf. D.G. Lupiáñez et al., 2015). Autrement dit, pour pouvoir exprimer un gène, certaines régions d'un compartiment doivent être proches les unes des autres afin de pouvoir s'agglutiner et former des amas de chromatine. Lorsque deux régions sont éloignées dans la séquence, il se forme alors une boucle de chromatine. Si plus de deux régions doivent être rapprochées, il se forme un amas plus complexe, un TAD. Grâce à la méthode Hi-C, les chercheurs peuvent obtenir une image de l'activité des TAD à un instant donné. De plus, grâce à des microscopies à l'échelle nanométrique (10-5 à 10-4 mm), les chercheurs ont pu cartographier de manière précise le mode de repliement de la chromatine à l'intérieur des TAD, révélant des nanodomaines de chromatine ou CND (cf. F.Bantignies et al., 2019). La découverte de ces différentes échelles d'organisation des chromosomes a permis de mieux comprendre le fonctionnement du génome, ainsi que des maladies dont l'origine restait mystérieuse. On sait par exemple que des réarrangements chromosomiques comme des inversions ou des délétions surviennent au niveau des régions en bordure des TAD. Cela entraîne des défauts d'isolement entre les gènes. Par conséquent, des séquences stimulatrices localisées dans le TAD voisin peuvent communiquer et activer de manière erronée des gènes dans des tissus où ils sont normalement inactifs. On a ainsi découvert que des gènes du développement sont directement impliqués dans des malformations des membres (Syndrome F - une anomalie squelettique -, la polydactylie, etc), des défauts neurologiques ou des dysplasies. Des réarrangements chromosomiques peuvent également entraîner l'expression de gènes normalement muets pouvant conduire à l'apparition de tumeurs. Grâce à ces travaux, on comprend mieux l'organisation spatiale du génome et on peut expliquer de nombreux "défauts" cellulaires liés à la dérégulation des gènes. Mais il reste de nombreuses questions sans réponses. Ainsi on ignore quel mécanisme détermine la taille et les bordures des TAD et des CND ou quel est le rôle des CND dans la régulation des fonctions du génome. Enfin, même en connaissant la séquence de l'ADN et son organisation tridimensionnelle, on ne peut toujours pas prédire dans un type cellulaire donné, comment son expression génique est régulée. Voilà autant de sujets de recherche pour les futurs thésars ou prix Nobel. Codons, bases et acides aminés Les acides aminés sont des substances fabriquées (codées) par les gènes présents dans les chromosomes. Ces substances sont indispensables au bon fonctionnement des processus métaboliques et physiologiques cellulaires et notamment au stockage de toutes les substances nutritives dont le corps a besoin : eau, protéines, lipides, oligo-éléments, vitamines. Autrement dit, leur déficience entraîne des carences et des maladies (fatigue, obésité, cholestérol, diabète, insomnie, perte des cheveux, accentuation des rides, etc). Tous les acides aminés ont une fonction acide (COOH) et amine (NH2) plus un radical, une molécule variable qui les différenciera. Le nombre d'acides aminés possible est lié à la combinaison des bases simples A, G, T, C ou U (uracile). Sur 64 combinaisons possibles, le code génétique ne code que 21 acides aminés dits protéinogènes. Le plus simple est la méthionine (Met) seule combinaison de bases ou codon (AUG), jusqu'à l’arginine (Agr) et la leucine (Leu) formées de 6 codons. Parmi eux la sélénocystéine (Sec) joue un rôle particulier car elle n'est pas à proprement parlé encodée par un codon mais dans l'ARNm, à la position correspondant à la sélénocystéine dans la protéine se trouve un codon stop opale UGA dont la lecture par le ribosome provoque l'arrêt de la traduction et donc de la synthèse protéique. Pour être complet, il existe un 22e acide aminé protéinogène, la pyrrolysine (Pyl), codée par le codon UAG mais il n'est exprimé que chez certains organismes archéens méthanogènes. A
voir : Comment fait-on une analyse
ADN ?, Unisciel Séquençage de l'ADN, Enième Prod Les acides aminés (PDF), Michel Sève
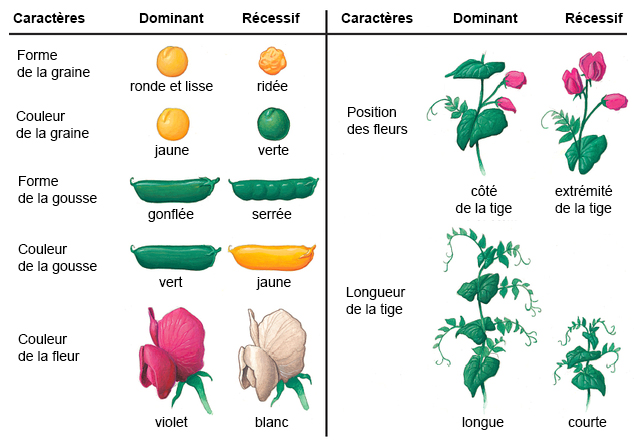
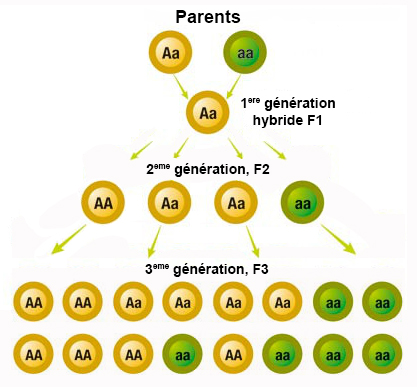
Nos 4 lettres qui forment l'alphabet de la vie nous donnent ainsi accès aux processus de reproduction et de contrôle des organismes vivants. C'est la mutation de cette structure simple qui aboutit au bout de 4.5 milliards d'années à l'ancêtre de l'Homme. Notons que dans la nature il existe au moins 500 acides aminés dont environ 149 existent sous forme de protéines, 22 acides aminés sont fondamentaux dont 19 ne comprennent que 4 éléments chimiques (C, H, O et N), deux autres contiennent également du soufre (la méthionine et l'homocystéine) et un troisième contient du sélénium (sélénocystéine). Génotype et phénotype Les lois de Mendel En réalisant des expériences de fécondation croisée avec des graines de pois notamment et en observant de quelle manière les plants et les pois changeaient de forme et de couleurs au fil des générations, en 1860 le moine et botaniste tchèque Grégor Mendel découvrit que les caractères héréditaires se répartissaient de façon précise chez les individus. Aujourd'hui on préciserait sa pensée en disant que les gènes se transmettent d'une génération à l'autre en exprimant des caractères dominants et récessifs. Les travaux de Mendel ne furent pas reconnus de son vivant mais il avait vu juste quand il disait "un jour mon heure viendra". Mendel avait en effet découvert les lois de l'hérédité, ce que ses successeurs appelleront les lois de Mendel qui se résument comme suit : 1. La loi d'uniformité des hybrides de première génération : si on croise deux variétés d'individus de race pures se distinguant par un seul caractère (homozygotes), tous les descendants de la première génération, les hybrides F1, sont identiques. Autrement dit, si les parents sont de souche pure, aucune forme intermédiaire n'apparaît à la première génération (en excluant les mutations). 2. La loi de disjonction des allèles : lors du croisement entre deux des individus de générations F1, c'est-à-dire des individus qui se différencient par 2 caractères, on obtient à la deuxième génération F2 quatre types distincts dans des proportions 3:1. Autrement dit, les facteurs héréditaires se séparent dans les gamètes, chaque gamète ne contenant qu'un facteur de chaque caractère. 3. La loi de ségrégation : le croisement des individus de générations F2 (possédant quatre génotypes distincts) formera toutes les combinaisons possible. Autrement dit la ségrégation est indépendante des caractères héréditaires multiples. Cette règle ne s'applique que si les gènes responsables des caractéristiques acquises se situent sur différents chromosomes ou s'ils sont éloignés sur le même chromosome. A
voir : Les
lois de Mendel Les génies de la génétique, Guilgamesh/La Cinquième
Le rôle des allèles Le caractère d'un individu dépend de son génotype, de l'expression des gènes (bien que l'environnement joue également un rôle). L'homme partage au moins 25% de ses gènes avec les autres créatures vivantes, des mammifères aux poissons en passant par les végétaux. Seule différence, ceux-ci sont simplement plus éloignés de nous dans le temps, leur génome s'étant modifié au cours de l'évolution. Chaque gène occupe une place précise sur un chromosome, ce qu'on appelle le locus. Cet emplacement génétique spécifique permet aux généticiens d'associer des caractéristiques biologiques à l'expression d'un gène. Mais tous ne sont pas exprimés.
Sur l'ensemble du génome humain, environ 60% soit ~12000 gènes sont exprimés et ce nombre à tendance à diminuer à mesure que les recherches s'affinent (cf. M.L. Tress et al., 2014). Il est important de distinguer le génotype d'un individu, c’est-à-dire son patrimoine génétique, et le phénotype, c'est-à-dire son apparence physique, ses traits ou caractères. Le caractère dominant ou récesssif de certains gènes portés par les chromosomes crée autant de versions différentes des gènes qu'on appelle les allèles. C'est leur variabilité qui explique les particularités de chaque individu tout en héritant des caractères généraux de l'espèce. Si les deux allèles sont identiques, l'individu est dit homozygote pour ce locus. Si les allèles sont différentes, l'individu est dit hétérozygote pour ce locus. Enfin, il y a le cas particulier des mâles hémizygotes, c'est-à-dire porteurs des gènes du chromosome X. A l'inverse, le phénotype décrit l'individu selon l'expression du génotype en réponse à la pression de l'environnement. Dans une population d'individus, il peut y avoir un grand nombre d'allèles d'un même gène sans pour autant qu'ils soient exprimés chez tous les individus. Ainsi, le gène OCA2 qui détermine la couleur des yeux chez l'être humain, peut avoir des allèles codant pour la couleur bleue, verte, noisette ou marron. La couleur marron est dominante tandis que le bleu est récessif (il ne produit un effet que s'il est présent sur les deux chromosomes appariés). Un individu possédant deux allèles normaux et un individu possédant un allèle normal et un allèle mutant présentent généralement le même phénotype. En revanche, un individu ayant deux allèles mutants présentera un phénotype différent d’un organisme possédant au moins un allèle normal. Mis à part le cas des mutations, le phénotype d'un individu peut changer au cours de sa vie alors que son génotype reste constant. Voyons à présent en détails certains mécanismes génétiques et cellulaires. Prochain chapitre Le sens de l'information génique
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||







{kind=link}